Impact of L-Arginine Supplementation on Thyroid Function in Male Wistar Rats Exposed to Dichlorvos
-
Saka Waidi Adeoye
Department of Physiology, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo State, Nigeria
Igbayilola, Yusuff Dimeji

Department of Human Physiology, College of Medicine and Health Sciences, Baze University, Abuja, Nigeria
Dada Kafilat OmolaraDepartment of Physiology, College of Health Sciences, Ladoke Akintola University of Technology, Ogbomoso, Oyo State, Nigeria
Grema Mariam GujjaDepartment of Human Physiology, College of Medicine and Health Sciences, Baze University, Abuja, Nigeria
| Received 05 May, 2024 |
Accepted 26 May, 2024 |
Published 27 May, 2024 |
Background and Objective: Thyroid hormones cause the body to burn oxygen and produce heat. Thyroid hormone has a wide range of functions, therefore imbalances or excesses in hormone levels result in a variety of clinical symptoms. The study looks into how supplementing with L-arginine affects thyroid function in male Wistar rats that have been exposed to the common herbicide Dichlorvos (DDVP). The investigation of possible treatment strategies is warranted since thyroid dysfunction is a common health concern linked to environmental contaminants. Materials and Methods: The four groups of male Wistar rats were treated with control, dichlorvos, L-arginine and dichlorvos with L-arginine supplementation. Following treatment, thyroid function markers such as Triiodothyronine (T3), Thyroxine (T4) and Thyroid-Stimulating Hormone (TSH) levels were measured. Results: The dichlorvos-exposed group’s thyroid hormone levels were significantly different from the controls’, suggesting thyroid malfunction, according to the results. On the other hand, rats given L-arginine supplements in addition to dichlorvos exposure demonstrated significant preservation of thyroid function as shown by somewhat normalized hormone levels. Conclusion: This implies that L-arginine may have a protective effect against thyroid disturbance brought on by dichlorvos. Human health may be affected by learning how L-arginine supplementation reduces pesticide-induced thyroid dysfunction, especially in groups of people who are more susceptible to exposure to environmental toxins.
| Copyright © 2024 Adeoye et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Thyroid hormones are calorie-producing, they cause the body to burn oxygen and produce heat. They boost gluconeogenesis, enhance glucose uptake, accelerate protein catabolism and advance lipid metabolism. Regarding additional effects specific to an organ, thyroid hormones affect cardiac function by decreasing peripheral vascular volume and increasing cardiac output, heart rate, myocardial contractility and blood volume1. To promote bone growth and development, they increase the synthesis of growth factors, cytokines and other substances2. Thyroid hormone has a wide range of functions, therefore imbalances or excesses in hormone levels result in a variety of clinical symptoms. Those who have related problems experience severe discomfort due to these shortages or elevations that affect several organ systems1.
It has been reported that target organs of dichlorvos toxicity include the Central Nervous System (CNS) and less in other organs of the body. This indirectly affects the levels of hormones affected by the hypothalamus3. Presently, the increasing health challenges involving the CNS call for a proper study on the chronic toxicological effects of dichlorvos using rabbits to carry out further studies in man. Animal studies have shown a decrease in spermatogenesis in relation to dichlorvos toxicity3. By harming the seminiferous epithelium through germ cell proliferation, other organophosphates such as parathion, methyl parathion and chloropyrifos can also have an impact on sperm counts3. Nonetheless, a reduction in LH levels may result from dichlorvos’ impact on hypothalamic-pituitary endocrine activities. Dichlorvos reduces brain AChE activity, which modifies hypothalamic-pituitary endocrine functioning. Changes in serum reproductive hormones, such as testosterone, have been observed in men who were exposed to dichlorvos4. A positive relationship between dichlorvos and total testosterone level has been observed in a cross-sectional study in male Thai5.
Food contains L-arginine, an endogenous amino acid that is safe to consume. According to Wu and Morris6, the molecule is involved in the synthesis of several products that carry out regulatory tasks within the body. Among these, nitric oxide is especially significant since it is a signaling molecule that controls the metabolism of fats and carbohydrates6. Although the mechanisms underlying these effects have not been sufficiently clarified, there is growing experimental and clinical data suggesting that L-arginine supplementation may be useful in managing disturbed metabolism in obesity, regulating arterial blood pressure, or easing the symptoms of type 2 diabetes6. The L-arginine is a semi-essential amino acid that is used by all cells6.
This amino acid is absorbed in the small intestine’s jejunum and ileum and makes up, on average, 5-7% of all the amino acids in a typical human diet. The body uses l-arginine for immune cell function, tissue healing, protein synthesis and the urea cycle6and supplementation with L-arginine has been shown to ameliorate the pathology of various diseases in clinical practice7. Citrulline, a vasodilator and nitric oxide are produced from arginine. Nitric oxide synthase (NOS) exists in three isoforms; to synthesize nitric oxide, these isoforms require oxygen, arginine, Tetrahydrobiopterin 4 (BH4) and NADPH (Nicotine Amide Adenine Di Nucleotides Phosphate)8. Dichlorvos, an organophosphate insecticide, is known to cause various adverse health effects, including thyroid dysfunction. The thyroid gland plays a crucial role in regulating metabolism, growth and development, among other physiological processes. Exposure to dichlorvos has been linked to alterations in thyroid hormone levels, potentially leading to thyroid disorders such as hypothyroidism or hyperthyroidism. The L-arginine, a semi-essential amino acid, has been suggested to have protective effects against certain environmental toxins due to its role in nitric oxide synthesis and antioxidant properties.
However, the potential impact of L-arginine supplementation on thyroid function in the context of dichlorvos exposure has not been extensively studied, especially in animal models. Understanding the effects of L-arginine supplementation on thyroid function in rats exposed to dichlorvos could provide valuable insights into the mechanisms underlying thyroid dysfunction induced by organophosphate pesticides and offer a potential therapeutic approach to mitigate such effects. The study aimed to look into how supplementing with L-arginine affects thyroid function in male Wistar rats that have been exposed to the common herbicide dichlorvos (DDVP).
MATERIALS AND METHODS
Animals: Forty healthy male Wistar rats weighing between 220-250 g, obtained from a private breeder, were selected for the study. The use of Wistar rats in research offers a balance between practical considerations, genetic stability and biological relevance, making them valuable models for a wide range of scientific investigations. These rats had not undergone any prior experimental interventions and were considered healthy based on the absence of stress or infection indicators. Before initiating the experiments, the rats were individually weighed after a period of acclimatization lasting two weeks. They were housed in well-ventilated plastic cages at the animal facility of the Department of Physiology, Faculty of Basic Medical Sciences, College of Health Sciences, LAUTECH, Ogbomoso, Oyo State. During the research, the animals were provided with both food and water. The study was carried out between February to April, 2023.
Arrive statement: All animal experiments comply with the ARRIVE guidelines and are carried out in accordance with the U.K. Animals (Scientific Procedures) Act, 1986 and Associated Guidelines, EU Directive 2010/63/EU for animal experiments and the National Research Council’s Guide for the Care and Use of Laboratory Animals.
Ethical approval: Ethical approval was sought and given to conduct this study from Ethics and Review Committee of the Faculty of Basic Medical Sciences, Ladoke Akintola University of Technology, Ogbomoso with a reference number FBMS/AECP/092/23.
L-Arginine supplementation
Dosage and administration: A low dose of 100 mg/kg was used to determine the dose administered to each animal9,10.
Chemical: A low dose of 100 mg/kg was used to calculate the dosage given to each animal. Based on the formula weight (g)×dosage (100 mg/1000 g), the computation was performed with an average weight of 250 g for the rats. The amount of L-arginine given was calculated based on the fact that one 500 mg capsule of L-arginine includes ten rats totaling 2500 g in weight. The proportion for 2500 g (x) of L-arginine was calculated using 1000 g containing 100 mg of L-arginine. As a result, the group of 10 male Wistar rats received 250 mL of L-arginine. Consequently, 25 mg of L-arginine were given to each rat10,11.
Determination of body weight: Rats were weighed before dosing and weekly during the experiment using a scale. The weight of the animal is necessary to determine the dose of medicine administered to the animal (Salter, England).
Grouping: They were divided into four groups of ten animals each as follows:
| • | Group A (control group): They received only food and water throughout the study | |
| • | Group B (only dichlorvos): This group was exposed to dichlorvos by inhalation for 10 min in a desiccator containing 2 mL of dichlorvos, which was moistened daily for three weeks without treatment | |
| • | Group C (dichlorvos (DDVP)+L-arginine): This group was exposed to dichlorvos in the same way as group B and treated with oral L-arginine for six weeks | |
| • | Group D (only L-arginine): This group received only one supplemental dose of L-arginine per day for six weeks |
Collection of blood samples: Blood sample collection occurred after an overnight fast12,13 at the conclusion of the experimental period. To euthanize the animals, ketamine was administered as a sedative. A vertical incision was promptly made in the abdomen and blood was obtained through cardiac puncture using a 2 mL needle and syringe, then transferred into standard vials14.
Determination of Triiodothyronine (T3) measurement: The level of Triiodothyronine (T3) concentration was determined by ELISA kit from Elabscience, Wuhan, China using rat T3 ELISA kit. The microplates’ wells were prepared for each serum reference, control and patient specimen, with duplicates for each assay. Any unused microwell strips were returned to the aluminum bag, sealed and stored at 2-8°C. Then, 0.050 mL (50 μL) of the appropriate serum reference, control, or specimen was pipetted into the assigned well. Following this, 0.100 mL (100 μL) of Working Tracer, T3-enzyme conjugate solution was added to all wells and the microplate was gently swirled for 20-30 sec to mix and cover. It was then incubated for 45 min at room temperature. The contents of the microplate were discarded either by decantation or aspiration and if decanting was used, the plate was blotted dry with absorbent paper. Subsequently, 350 μL of wash buffer was added and decanted (tap and blot) or aspirated, with this process repeated four additional times for a total of five washes. An automatic or manual plate washer could be used, following the manufacturer’s instructions. If a squeeze bottle was used, each well would have been filled by depressing the container (to avoid air bubbles) to dispense the wash, which was then decanted, with this process being repeated four additional times. Following this, 0.100 mL (100 μL) of working signal reagent solution was added to all wells, adhering to the same order of reagent addition to minimize reaction time differences between wells. The microplate was then incubated for 5 min in the dark at room temperature. Finally, the relative light units in each well were read for a minimum of 0.5-1.0 sec, using a microplate luminometer and the results were read within thirty minutes of adding the signal solution. Using a reference wavelength of 620-630 nm to reduce well defects, the absorbance in each well was measured at 450 nm using a microplate reader from Elabscience, Wuhan, China using.
Thyroxine (T4) measurement: The serum T4 level was determined using the Enzyme-Linked Immunosorbent Assay (ELISA) test, employing a rat T4 ELISA kit from Elabscience, Wuhan, China. The microplate wells were formatted for each serum reference, control and patient specimen, with duplicates for each assay. Any unused microwell strips were replaced into the aluminum bag, sealed and stored at 2-8°C. Then, 0.050 mL (50 μL) of the appropriate serum reference, control or specimen was pipetted into the assigned well. Following this, 0.100 mL (100 μL) of T4 enzyme reagent was added to all wells and the microplate was gently swirled for 20-30 sec to mix and cover. It was then incubated for 60 min at room temperature. The contents of the microplate were discarded either by decantation or aspiration and if decanting was used, the plate was blotted dry with absorbent paper. Subsequently, 350 μL of wash buffer was added and decanted (tap and blot) or aspirated, with this process repeated two additional times for a total of three washes. A plate washer from Elabscience, Wuhan, China was used, following the manufacturer’s instructions. If a squeeze bottle was employed, each well was filled by depressing the container (avoiding air bubbles) to dispense the wash, which was then decanted, with this process repeated two additional times. To reduce variations in reaction times between wells, 0.100 mL (100 μL) of working substrate solution was then added to each well in the same order as the reagents. After that, the microplate was incubated for 15 min at room temperature. Lastly, each well received 0.050 mL (50 μL) of stop solution, which was carefully stirred for 15 to 20 sec. Using a reference wavelength of 620-630 nm to reduce well defects, the absorbance in each well was measured at 450 nm using a microplate reader.
Thyroid Stimulating Hormone (TSH) measurement: The serum TSH level was determined using the Enzyme-Linked Immunosorbent Assay (ELISA) test, employing a rat T4 ELISA kit from Elabscience, Wuhan, China. Appropriate number of coated strips were put in the holder. Fifty milliliters of TSH standards, control and specimens were pipetted into the appropriate wells. One hundred milliliters of the conjugate reagent, ready for use, was added to each well. For 10 to 30 sec, shook. placed a cover on the plate and let it sit at room temperature (18-26°C) for 60 min. Liquid was extracted from every well. Three times, 300 mL of 1x wash buffer was used to wash the wells. Spread out over absorbing paper towels. The TMB substrate (100 mL) was added to each well. Incubated at room temperature for 15 min. Fifty milliliters of stop solution was added to each well. To combine the solution, the plate was gently shaken. After adding the stopping solution, wait 15 min and check the absorbance using an ELISA reader set to 450 nm.
Statistical analysis: Data were recorded as Mean±Standard Error of the mean. Statistical difference between the means was determined by One-way ANOVA. The Tukeys’ post hoc test is used to identify differences between individual means. The confidence interval was set at 95%, so that in all cases results with a value of p<0.05 were accepted as being significant (GraphPad Prism 5, GraphPad Software, Inc., La Jolla, California, USA).
RESULTS
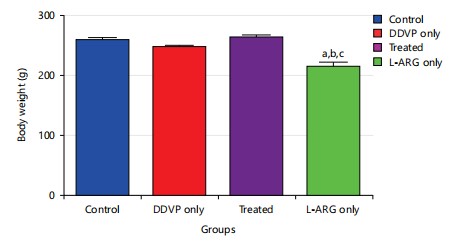
Effect of L-arginine on body weight changes in DDVP-exposed rats: The results (Fig. 1) revealed that exposure of animals to dichlorvos caused a non-significant change in the weight of the animals when compared to control at p>0.05. Also, administration of L-arginine caused a non-significant increase in body weight of the rats when compared to DDVP-exposed at p>0.05.
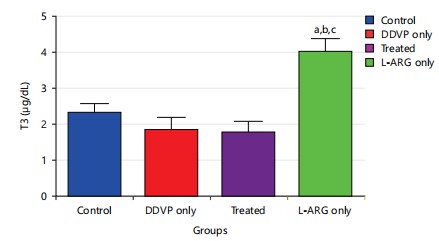
Effect of L-arginine on Triiodothyronine (T3) in DDVP-exposed rats: The results (Fig. 2) revealed that exposure of animals to dichlorvos caused a non-significant change (p>0.05) in Triiodothyronine (T3) levels when compared to control. Also, administration of L-arginine caused a non-significant change p>0.05 in Triiodothyronine (T3) levels when compared to DDVP-exposed rats. However, rats exposed to L-arginine had a significant increase p<0.05 in T3 levels compared with control, DDVP only and DDVP+L-arginine rats.
|
|
|
|
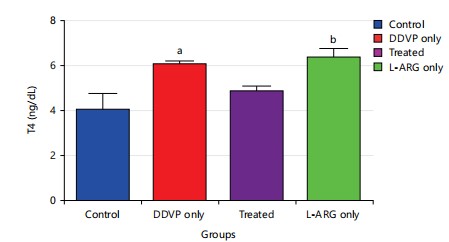
Effect of L-arginine on Thyroxine (T4) in DDVP-exposed rats of the animals: The results (Fig. 3) revealed that exposure of animals to dichlorvos caused a significant increase p<0.05 in Thyroxine (T4) levels when compared to control. Also, administration of L-arginine caused a non-significant decrease in Thyroxine (T4) levels when compared to DDVP-exposed at p>0.05. In addition, rats exposed to L-arginine only had a significant increase p>0.05 in Thyroxine (T4) levels compared with control, DDVP only and DDVP+L-arginine rats.
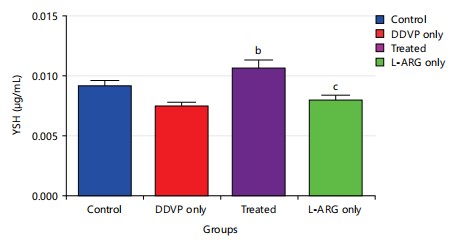
Effect of L-arginine on Thyroid Stimulating Hormone (TSH) in DDVP-exposed rats of the animals: The results (Fig. 4) revealed that exposure of animals to dichlorvos caused a non-significant decrease p>0.05 in thyroid-stimulating hormones when compared to control. Also, administration of L-arginine caused a significant increase p>0.05 in thyroid-stimulating hormone levels when compared to DDVP-exposed rats.
DISCUSSION
This study was carried out to investigate the effects of l-arginine supplementation on thyroid function parameters of dichlorvos-exposed male Wistar rats. It has been reported that target organs of dichlorvos toxicity include the Central Nervous System (CNS) and less in other organs of the body. This indirectly affects the levels of hormones affected by the hypothalamus3. Thyrotropin-Releasing Hormone (TRH), a tripeptide secreted into the venous system that drains to the pituitary gland, is produced by the hypothalamus12. Thyrotropin or Thyroid-Stimulating Hormone (TSH), is produced and secreted by the pituitary gland when Thyroid-Stimulating Hormone (TRH) attaches to receptors in thyrotroph cells. Thyroid stimulating hormone stimulates the production and secretion of thyroid hormones, T3 and T4, by binding to TSH receptors in the thyroid gland’s follicular cells.
The TSH is a glycoprotein that interacts with FSH receptors to cause precocious puberty, also known as Van Wyk-Grumbach syndrome13. It shares its alpha subunit with LH and FSH. Leydig cells and human follicular cells have also been shown to carry TSH receptors15,16. Moreover, TSH induces endometrial tissue proliferation in endometriosis-affected women17. Furthermore, some evidence suggests that TSH influences leptin’s ability to regulate kisspeptin-neurokinin B-dynorphin neurons18,19. This is because TSH activates TSHR, which in turn affects lipolysis, adipocyte and preadipocyte proliferation and leptin secretion20,21. Disorders of the thyroid gland are usually described by the level of circulating hormone which includes hypothyroidism, hyperthyroidism, euthyroid sick syndrome and those resulting from medications22. The present study revealed that exposure to dichlorvos caused a significant decrease in the level of Thyroid Stimulating Hormone (TSH) which is suggestive of thyroid dysfunction. Reduction in the levels of T3 is seen in this study which might be a result of structural damage of thyroid follicular cells due to accumulation of dichlorvos in the thyroid gland and its effects on the regulatory enzymes associated with hypothalamic pituitary thyroid (HPT) axis, this result was in agreement with the study done by Ross et al.23. Increase in the level of tetrathyroxine is seen in Dichlorvos-exposed rats when compared with control rats.
Although the body can synthesize L-arginine on its own, it has also been demonstrated that exogenous intake through diet supplements can augment the body’s supply and address changes in metabolism. According to Borger et al.24, it has been shown to be the essential substrate in the nitric oxide (NO) pathway that inhibits the renin-angiotensin system (RAS) and partially regulates insulin release. When L-arginine was administered instead of dichlorvos-exposed individuals, an increase in thyroid stimulating hormone and triiodothyronine was observed. In contrast to rats exposed to dichlorvos, treated rats exhibit lower thyroxine levels.
Nevertheless, it is important to recognize a number of the current study’s limitations in spite of the encouraging outcomes. First of all, the research used an animal model, which might not accurately represent the intricacies of pesticide exposure and thyroid physiology in humans. Furthermore, this study did not fully clarify the precise processes underlying the protective effects of L-arginine supplementation, which calls for additional research. Future studies should use both in vitro and in vivo models to clarify the molecular mechanisms behind L-arginine-mediated protection against pesticide-induced thyroid dysfunction.
CONCLUSION
The current research shows that in male Wistar rats exposed to dichlorvos, thyroid function is significantly preserved by L-arginine supplementation. The results point to a possible therapeutic approach using L-arginine to mitigate thyroid dysfunction caused by pesticides. To fully understand the underlying mechanisms and evaluate the translational potential of supplementing with L-arginine to prevent thyroid disorders caused by environmental toxins, more research is necessary. Overall, it is proposed that L-arginine supplementation will exert protective effects on thyroid function in male Wistar rats exposed to dichlorvos, potentially offering a novel therapeutic strategy for mitigating pesticide-induced thyroid dysfunction.
SIGNIFICANCE STATEMENT
This work is significant because it looks at how supplementing with L-arginine affects thyroid function in male Wistar rats that have been exposed to dichlorvos. Dichlorvos is a widely used pesticide that has been connected to thyroid dysfunction, which is dangerous for both environmental and human health. Second, L-arginine appears to be a promising agent in combating the effects of toxins due to its demonstrated vasodilatory and antioxidant qualities. Its potential to guard against thyroid disruption caused by dichlorvos may help prevent or treat similar toxin exposures. Finally, gender-specific response analysis can be carried out by focusing on male Wistar rats, enhancing the understanding of susceptibility variations. Overall, this work adds to a larger body of environmental research.
REFERENCES
- Talat, A., A.A. Khan, S. Nasreen and J.A. Wass, 2019. Thyroid screening during early pregnancy and the need for trimester specific reference ranges: A cross-sectional study in Lahore, Pakistan. Cureus, 11.
- Zhu, B., G. Zhao, L. Yang and B. Zhou, 2018. Tetrabromobisphenol a caused neurodevelopmental toxicity via disrupting thyroid hormones in zebrafish larvae. Chemosphere, 197: 353-361.
- Babazadeh, M. and G. Najafi, 2017. Effect of chlorpyrifos on sperm characteristics and testicular tissue changes in adult male rats. Vet. Res. Forum, 8: 319-326.
- Melgarejo, M., J. Mendiola, H.M. Koch, M. Moñino-García, J.A. Noguera-Velasco and A.M. Torres-Cantero, 2015. Associations between urinary organophosphate pesticide metabolite levels and reproductive parameters in men from an infertility clinic. Environ. Res., 137: 292-298.
- Neghab, M., M. Momenbella-Fard, R. Naziaghdam, N. Salahshour, M. Kazemi and H. Alipour, 2014. The effects of exposure to pesticides on the fecundity status of farm workers resident in a rural region of Fars Province, Southern Iran. Asian Pac. J. Trop. Biomed., 4: 324-328.
- Wu, G. and S.M. Morris Jr., 1998. Arginine metabolism: Nitric oxide and beyond. Biochem. J., 336: 1-17.
- Rashid, J., S.S. Kumar, K.M. Job, X. Liu, C.D. Fike and C.M.T. Sherwin, 2020. Therapeutic potential of citrulline as an arginine supplement: A clinical pharmacology review. Pediatr. Drugs, 22: 279-293.
- White, M.F., 1985. The transport of cationic amino acids across the plasma membrane of mammalian cells. Biochim. Biophys. Acta-Rev. Biomembr., 822: 355-374.
- Ramos, L., R. Labat, F.A.S. Carvalho, A.B. Martin and R.Á.B. Lopes-Martins, 2006. Effect of oral arginine administration over blood pressure and cardiac parameters in rats submitted to chronic inhibition of nitric oxide synthesis. Rev. Bras. Med. Esporte, 12: 169-174.
- Saka, W.A., T.E. Ayoade, T.M. Akhigbe and R.E. Akhigbe, 2021. Moringa oleifera seed oil partially abrogates 2,3-dichlorovinyl dimethyl phosphate (Dichlorvos)-induced cardiac injury in rats: Evidence for the role of oxidative stress. J. Basic Clin. Physiol. Pharmacol., 32: 237-246.
- Nwankwo, R.C., M.D. Ibegbu, K.C. Onyekwelu, C.S. Ejezie, J.E. Ikekpeazu and F.E. Ejezie, 2019. Biochemical and histopathological effects of sub-acute exposure of albino rats to fumigants-dichlorvos and cypermethrin. Interdiscip. Toxicol., 12: 180-185.
- Igbayilola, Y., A. Morakinyo and B. Iranloye, 2021. Leptin-resistance induced hyperphagia and diminished oxidative balance in offspring of dams exposed to perinatal protein restriction. Afr. J. Biomed. Res., 24: 451-458.
- Dimeji, I.Y., A.O. Samson, A. Kayode, A. Ikponwosa, W.O. Dada and O.F. Adesina, 2023. Glucometabolic response to walnut (Juglans regia L.) supplementation during gestation and/or lactation in offspring of Sprague-Dawley rats. Singapore J. Sci. Res., 13: 79-87.
- Igbayilola, Y.D. and M.G. Gujja, 2024. Alpha-amylase and alpha-glucosidase upregulated glucose homeostasis in high-fat fed wistar rats supplemented with cocoa flavonoid-rich aqueous. Food Biosci., 59.
- USPSTF, 2004. Screening for thyroid disease: Recommendation statement. Ann. Intern. Med., 140: 125-127.
- Anasti, J.N., M.R. Flack, J. Froehlich, L.M. Nelson and B.C. Nisula, 1995. A potential novel mechanism for precocious puberty in juvenile hypothyroidism. J. Clin. Endocrinol. Metab., 80: 276-279.
- Dhole, B., S. Gupta, S. Shekhar and A. Kumar, 2020. A novel antigonadotropic role of thyroid stimulating hormone on leydig cell-derived mouse leydig tumor cells-1 line. Ann. Nat. Acad. Med. Sci., 56: 30-37.
- Agard, J.A., D.M. Duffy, T. Jacot and D.F. Archer, 2011. Thyroid stimulating hormone (TSH) receptor on granulosa cells. Fertil. Sterility, 96.
- Peyneau, M., N. Kavian, S. Chouzenoux, C. Nicco and M. Jeljeli et al., 2019. Role of thyroid dysimmunity and thyroid hormones in endometriosis. Proc. Natl. Acad. Sci. U.S.A., 116: 11894-11899.
- Christen, T., S. Trompet, R. Noordam, J.B. van Klinken and K.W. van Dijk et al., 2018. Sex differences in body fat distribution are related to sex differences in serum leptin and adiponectin. Peptides, 107: 25-31.
- Petrine, J.C.P., C.R. Franci and B.D. Bianco-Borges, 2020. Leptin actions through the nitrergic system to modulate the hypothalamic expression of the kiss1 mRNA in the female rat. Brain Res., 1728.
- Davies, T., R. Marians and R. Latif, 2002. The TSH receptor reveals itself. J. Clin. Invest., 110: 161-164.
- Ross, D.S., H.B. Burch, D.S. Cooper, M.C. Greenlee and P. Laurberg et al., 2016. 2016 American thyroid association guidelines for diagnosis and management of hyperthyroidism and other causes of thyrotoxicosis. Thyroid, 26: 1343-1421.
- Böger, R.H., S.H. Bode-Böger, R.P. Brandes, L. Phivthong-Ngam and M. Böhme et al., 1997. Dietary L-arginine reduces the progression of atherosclerosis in cholesterol-fed rabbits: Comparison with lovastatin. Circulation, 96: 1282-1290.
How to Cite this paper?
APA-7 Style
Adeoye,
S.W., Dimeji,
I.Y., Omolara,
D.K., Gujja,
G.M. (2024). Impact of L-Arginine Supplementation on Thyroid Function in Male Wistar Rats Exposed to Dichlorvos. Trends in Medical Research, 19(1), 151-159. https://doi.org/10.3923/tmr.2024.151.159
ACS Style
Adeoye,
S.W.; Dimeji,
I.Y.; Omolara,
D.K.; Gujja,
G.M. Impact of L-Arginine Supplementation on Thyroid Function in Male Wistar Rats Exposed to Dichlorvos. Trends Med. Res 2024, 19, 151-159. https://doi.org/10.3923/tmr.2024.151.159
AMA Style
Adeoye
SW, Dimeji
IY, Omolara
DK, Gujja
GM. Impact of L-Arginine Supplementation on Thyroid Function in Male Wistar Rats Exposed to Dichlorvos. Trends in Medical Research. 2024; 19(1): 151-159. https://doi.org/10.3923/tmr.2024.151.159
Chicago/Turabian Style
Adeoye, Saka, Waidi, Igbayilola, Yusuff Dimeji, Dada Kafilat Omolara, and Grema Mariam Gujja.
2024. "Impact of L-Arginine Supplementation on Thyroid Function in Male Wistar Rats Exposed to Dichlorvos" Trends in Medical Research 19, no. 1: 151-159. https://doi.org/10.3923/tmr.2024.151.159

This work is licensed under a Creative Commons Attribution 4.0 International License.